Regional emperor penguin population declines exceed modelled projections
June 10, 2025
Abstract
Emperor penguin populations are predicted to decline rapidly over the current century owing to habitat loss in Antarctica arising from warming oceans and loss of seasonal sea ice. Previous work using very high-resolution satellite imagery from 2009 to 2018 revealed a population decrease of 9.5%, characterized by a continuous decline until 2016, with a slight recovery until 2018. Our study, for the sector 0° to 90°W, includes the recent period of sea-ice loss between 2020 and 2023 and provides a regional population update for around a third of the global population. We used supervised classification of very high-resolution imagery, linked to a Markov model and Bayesian statistics. Results indicate a significant reduction in emperor penguin numbers, variance in the methodology is relatively high, but provides a best fit estimate of 22% decline over the period equating to a reduction of 1.6% per year. This decline exceeds the predictions of demographic models based on high-emission scenarios. It is unclear whether the sector analyzed here reflects conditions around the entire continent and our results highlight the need to extend the analysis to all sectors of Antarctica to determine whether these trends are reflected elsewhere.
Introduction
Stage-structured, process-oriented metapopulation demographic models1 suggest emperor penguin numbers will decline significantly by the end of this century, with almost all breeding sites extinct by 2100 under high greenhouse gas emission scenarios2. These models utilise Intergovernmental Panel on Climate Change (IPCC) climate predictions and sea-ice extent projection models; sea-ice extent3 together with land-fast ice extent4, are both known to be important factors in emperor penguin breeding success and long-term demographic processes2,5.
Over the last two decades, Antarctic sea-ice decline has not followed IPCC projections, where most models project a steady decline in sea ice extent6. The decline has been non-linear, with rapid loss in recent years7,8 and is now exhibiting record low levels9. Moreover, sea-ice extent, although easily measured, is not directly comparable to fast ice on which emperor penguins breed; therefore, sea-ice extent may not be fully representative of fast ice and short-term breeding success10. Nevertheless, in the long term the loss of large-scale sea ice in general means that ultimately, fast ice is likely to be affected. Years of low breeding success due to sea ice failure are critical for emperor penguin populations. Many other seabirds can abstain from breeding when conditions are poor, but this is not an option for adult emperor penguins who have been shown to breed almost every year11. Emperor penguins only produce one egg per season and have low juvenile survival rates12, therefore, multiple years of poor breeding success can be critical to population trends. With the projected future of the species at risk, there have been several recent attempts to raise the protection level of emperor penguins in Antarctica. In 2022, emperor penguins were listed under the U.S. Endangered Species Act Protection (26 October 2022: FWS-HQ-ES-2021-0043), whilst there have been proposals at the Antarctic Treaty Consultative Meeting (ATCM) to designate emperors an ‘Antarctic Specially Protected Species’ (ATCM, 2022)13. However, some Members of ATCM have posited that there is a lack of evidence to confirm that observed population trajectories are congruent with demographic model projections.
Emperor penguins are the only warm-blooded species to breed in the Antarctic during winter14. They breed around almost the whole Antarctic coastline, with the exception of the north-west Antarctic Peninsula, where fast ice is less stable4. Their breeding cycle, first described over 70 years ago15,16,17 makes the species extremely difficult to study using traditional methods18,19,20,21. Ship access to their breeding sites is often impossible in winter, whilst aerial surveys are operationally challenging. It is only over the last 15 years with the advent of satellite imaging22,23, that population assessments have been feasible at the circumpolar scale24. Regular ground counts have only been conducted at a small number of breeding sites and only a few colonies have long-term records (e.g. Pointe Géologie for multiple decades and shorter periods of monitoring for Atka Bay and several colonies in East Antarctica). However, single-site surveys are of limited use to study populations as movement between colonies, environmental variation and/or difference in age structure may vary between populations leading to different trends25,26. Therefore, a more regional approach must be taken27. The majority of the 66 known breeding sites have never had ground visits, many of them have never had aerial overflights and the populations at most of them are poorly known28,29.
The first global population estimate of 238,000 breeding pairs was determined from satellite images using a multivariate supervised classification approach24. As more colonies were discovered (e.g. Fretwell and Trathan28), revised population estimates of 258,000 pairs were identified2. Subsequently, a recent study assessed the populations of almost all emperor penguin colonies using a similar multivariate supervised classification supplemented by a Bayesian state-space population model30. The study covered a 10-year period between 2009 and 2018, finding a 9.5% global population decrease over that time. LaRue et al. 30 also considered the extent of sea-ice and fast ice and their connections to regional population trends, although results were inconclusive and trends in fast ice extent were only weakly correlated with the probability of population decline.
Importantly, during the period of the study by LaRue et al. 30, sea-ice extent around most of Antarctica was expanding31 until a significant change in 2016. Further, as sea-ice conditions have declined since 2018, it is uncertain how the emperor penguin population has responded in recent years. In particular, three consecutive years of record low spring sea-ice extent have occurred in 2022, 2023 and 2024 (https://nsidc.org/data/seaice_index). Associated with these years there have been catastrophic breeding failures at several colonies32,33, with relocation of breeding at multiple sites34.
Here we provide an updated population trend for the sector of Antarctica between 0° and 90° W, between Dronning Maud Land in the east and the Bellingshausen Sea in the west, an area that accounts for a third of the total population of the species30. Parts of this sector have experienced strong warming, with an increase in air temperatures of 2.8 °C between 1951 and 200035. Other parts of the sector have also shown change, with record low sea-ice in the Bellingshausen Sea36, and El Niño induced sea-ice changes around the Brunt Ice shelf33, both possibly affecting penguin populations. We use a similar multivariate supervised classification and statistical model to LaRue et al. 30, but extend the record for a further 5 years, from 2009 to 2023, thus capturing the more recent anomalous period of sea ice decline.
Methods and materials
Geographical area
The study area encompasses 0°–90° W (Fig. 1) and includes 19 emperor penguin colonies32, plus one colony believed to be functionally extinct37. The environment in this region spans the whole range experienced by emperor penguins elsewhere, from the marginal, fast-changing areas of the West Antarctic Peninsula and Bellingshausen Sea, to areas less affected by climate change, such as Dronning Maud Land and the Weddell Sea. Several colonies in our study area were only discovered after our study period commenced in 2009, and three of them have insufficient archival satellite imagery with which to estimate a population trend. Consequently, here we assess the trajectories of 16 colonies. The excluded sites were Gipps Ice Rise, Verdi Inlet and Cape Darlington; two of which (Gipps and Darlington) have small populations estimated to be below 500 birds, whilst Verdi Inlet is estimated to have around 3000 pairs33.
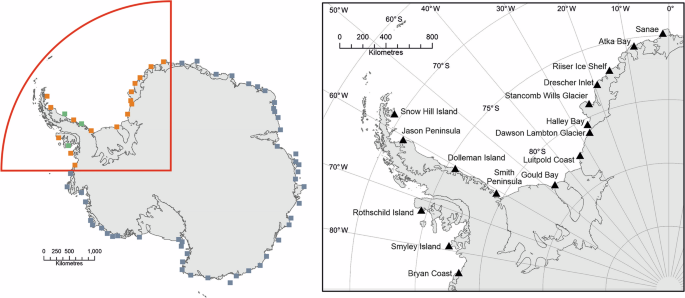
In the left panel orange squares show the 16 sites included in this study, green squares are sites with insufficient data, and grey squares represent the colonies outside our sector of interest. In the right panel, we identify the site names for the colonies included in this study.
Satellite imagery
We used optical satellite data from the MAXAR VHR satellite constellation (https://resources.maxar.com/brochures/the-maxar-constellation) that was speculatively tasked over each colony location each year, accessed from the MAXAR search and Discovery platform (https://discover.maxar.com/ and similar previous versions). A single section of each image with a minimum window of 25 km2 was obtained for each colony each year. In several cases, images were unsuitable as there was low cloud, low sun angle or poor environmental conditions obscuring or making penguins difficult to identify. Where possible, additional images were acquired. Additional information of image ID and scene quality is contained in Supplementary Table 5.
The analysis was restricted to the austral spring, between the end of August, when the sun comes up at the more northerly colonies, and the end of November before the adults depart the colonies as chicks fledge. The general method of pre-processing followed that of LaRue et al.30, first loading each image into ArcGIS (ESRI ArcGIS version 10.6 2024 and earlier versions), and projecting the images into the local UTM projection for more accurate area assessment. Imagery was pan-sharpened using ArcGIS, and enhanced using a ‘Standard Deviation’ histogram stretch. The location of the colony was then isolated, and the colony was cropped from the surrounding image manually to avoid excessive processing time. A supervised classification analysis was then conducted on each image, using a multivariate classification analysis in ArcGIS, which involves training a model on manually chosen pixels of penguin, guano, snow and shadow. These training data were chosen manually by experienced observers and usually equated to between 50-200 examples from each class, depending upon the homogeneity of the background image. Once the classes have been identified the multivariate classification algorithm divides the image into the available classes, one of which will be penguins. This process is iterative and usually training data needs to be refined multiple times before an acceptable confidence (>95% agreement between manual and automated observations) is reached. From these results, the penguin pixels were isolated and converted into a shapefile. The shapefile gives the area occupied by penguins in each image. This process was undertaken for a total of 241 images over the 15-year period (see Supplementary Data 5).
Statistical analysis
To convert the area of penguins in each image to an index of abundance, we followed an approach similar to that used by LaRue et al. 30, using a state-space analysis of emperor penguin population dynamics and observation process, and modified for satellite observations but without additional aerial or ground counts.
The population process accounted for daily changes in satellite counts over the survey period (August–November) in each of 16 identified colonies (Fig. 1). Colony-level trends and annual fluctuations were assessed considering the persistence of colonies at the same locations given physical changes (e.g. fast ice conditions), and occupancy (i.e. presence or absence for unknown reasons). The observation process accommodated bias and precision in satellite image counts due to data collection, and changes over survey period due to chick mortality and subsequent emigration by attendant and non-attendant adults.
Following LaRue et al. 30, expected abundance at colony j and year y was (N_j,y=z_j,yX_j,y), where (X_j,y) and (z_j,y) are terms for abundance and a colony presence. The inter-annual change in colony abundance ((r_j)) was modelled as (X_j,y sim Lognormal(log (X_j,y-1)+r_j,sigma _pro^2)), where (sigma _pro^2) is the process variance. Due to a relatively low sample size and to the sparse data of some colonies, colony-level effects were modelled as random effects with (r_j sim Normalleft(barr,sigma _r^2right)).
As in most state-space population models38, priors for initial population states ((X_j,1)) were given, and here we assumed a log-normal distribution with hyperparameters estimated empirically from the data as
Occupancy, or colony presence, was modelled with probability parameter p, as (z_j,y sim Bernoullileft(pright)). Although it was desired30, due to a low sample of years with colony absences, occupancy was not modelled accounting for detectability, and variation due to colony or previous occupancy (autocorrelation) effects.
The observation process assumed that the size of the areas occupied by penguins in satellite images ((N_j,y)) were normally distributed, centred on the expected number of penguins present in the colony on the day of the survey ((D_j,i,y)), and corrected for an estimated bias (beta _i) in interpretation of satellite images; (N_j,yexp (alpha D_j,i,y)beta _i). As the number of adults declines over the spring survey period, we used a log-linear fixed effect ((alpha)) of (D_j,i,y), which was the calculated day of the year corresponding to survey date and subtracting the average mid-point of the annual surveys (day 300). (alpha) thus described the proportional change in expected count for each day elapsed in the survey30.
Satellite counts were mapped to colony abundance as (S_j,i,y sim normal(N_j,yexp (alpha D_j,i,y)beta _i,w_j,y^2)), where (w_j,y^2=[(N_j,yexp (alpha D_j,i,y)beta _i)CV_S]^2). The variance term (w_j,y^2) assumed a constant coefficient of variation ((CV_S)), to be estimated when fitting the model, and assumed an increasing, but proportional, absolute error with colony size, which is common in image counts. Thus, separate fixed effects for (CV_S) and (beta _i) were estimated for each image quality score, which was arbitrarily assigned as poor (1), moderate (2) or good (3). These scores were related to the observers’ subjective ability to identify penguins in an image (similarly rated between 1 and 3), and to measured values of spectral band count, resolution, sun elevation, cloud cover, off nadir angle, collected GSD (ground sampling distance), and product GSD.
To accommodate observer bias, image quality scores were related to these variables in a discriminant analysis of principal components39, using R package adegenet40. Here, we selected the optimum number of principal components using cross-validation and used observer-assigned image quality as grouping variable in a discriminant analysis. The results provided a covariate with predicted image quality, and the subsequently fitted β coefficients assumed that satellite observations had a constant proportional bias.
Model fitting and assessment
We used Markov Chain Monte Carlo methods in program JAGS41, run from R (v.4.4.0; R Core Team, 2024)42 using package jagsUI43 to fit the state-space models. We used 550,000 iterations of three Markov chains using dispersed parameter values as starting values and discarded the first 50,000 samples of each chain as burn-in, thinning the remainder to every 50th sample, which produced 10,000 posterior samples from each Markov chain. We assessed chain convergence visually using trace plots, through the mixing of the chains and sample autocorrelation plots and using the (widehatR) potential scale reduction factor statistic44. We selected similar prior distributions as LaRue et al.30 (Supplementary Tables 1–3), and we assessed the model’s goodness-of-fit using posterior predictive checks. We obtained the root mean square error of simulated data and the observed data and obtained and estimated Bayesian p-value. P-values close to 0.5 indicated a reasonable fit and occurred when the fitted statistical model was equivalent to the ‘true’ model, which generated the data (Supplementary Fig. S1).
Results
Colony specific and general results can be seen in Fig. 2 and Supplementary Data 1–4.
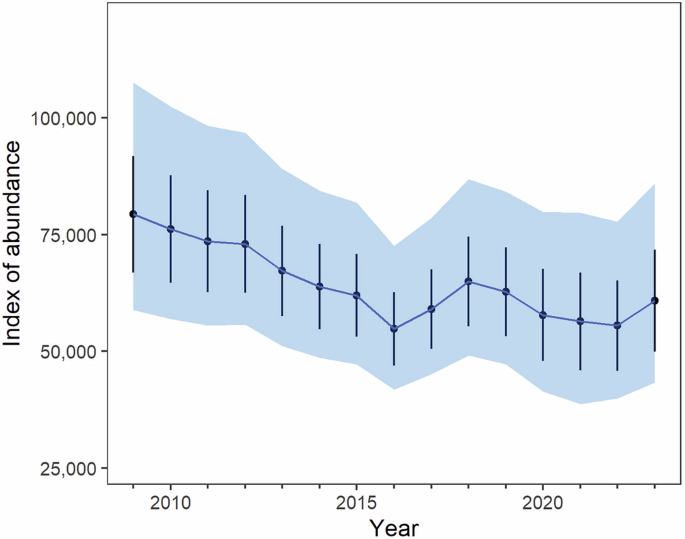
The blue area represents the 95% credible intervals and the solid value line the median of the posterior distribution simulations (in practical terms, these are the 0.025 and 0.975 percentiles of the posterior distribution of the abundance estimates). Black error bars are the mean estimates (or point estimates) of the abundance index plus and minus 1 standard error. The thicker blue line is the most likely scenario.
Estimates for model parameters describing the emperor penguin population dynamics are in Supplementary Table S1. All the (widehatR) values were lower than 1.1, and together with their trace plots suggested a successful convergence. Of the datasets simulated for posterior predictive checks, 24% had satellite counts with lower root mean square error (RMSE) than the observed data (Bayesian p-value = 0.24), suggesting a reasonable fit (Supplementary Fig. S1).
In the entire study area between 2009 and 2023 there was a 0.94 probability of emperor penguin population decline; a 0.30 probability of a 30% decline; and a 0.005 probability of a 50% decline. This corresponds to a likely mean decrease of −22% (95% CI: −44.6 to +8.34%) (Fig. 2), equivalent to approximately −18,275 fewer adults in 2023 (95% CI: −42415 to 5844) than in 2009.
The population trajectory (log-linear annual rate of change) was −2.25% per year (−4.33% to +0.04%), and an estimated abundance index of 59,400 (95% CI: 43,424–86,200) of penguin area in late spring, without an unknown quantity of pre-breeders, failed breeders, and non-breeders. The probability of a 30% decline over three emperor penguin generations (16 years on average) was estimated as 0.91.
Regional trends are summarised in Fig. 3, Table S2 and percent population change over the study period in Table S3. All of the regions except the Antarctic Peninsula showed a decline over the period of the study. Although the estimates of long-term abundance shown were highly variable, the brevity of the study period limited the ability to detect significant trends.
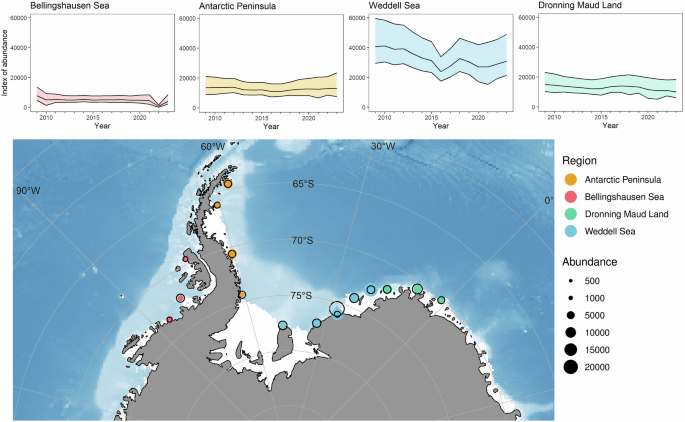
On the map, the size of the solid coloured circles denotes the relative size of the colony in 2023, the black outlines represent the size in 2009.
Discussion
Accuracy of the method
A primary concern when monitoring emperor penguins using VHR satellite imagery is the accuracy of the classification method. Uncertainty comes from two sources: (1) the errors associated with classifying the area of penguins as a metric of population; and (2) the potential variation in the population in late spring when the imagery is acquired45,46 The levels of variability from each of these two sources are potentially important45. With respect to the first issue, LaRue et al.30 and this study use images linked to Bayesian statistics to produce an overall population trend. Bayesian statistics are based on the concept that probability is a measure of belief in an event and gives confidence values based upon not just the overall probability, but the amount of data used to calculate those statistics (La Rue et al. analysed 460 images30, here we use 241). The population trend for both the LaRue et al.30 and our analysis is based upon methods that have been developed and refined by different groups over many years. The common practices that have evolved are considered robust and best practice for the data available. With respect to the second issue, it is important to recognise that the timing of image acquisition in late spring, dictates that only a proportion of the adult population is in attendance at each colony, with an unknown proportion at sea foraging, or having already failed in their breeding attempt. As such, it is uncertain whether the area classified as penguins in the images reflects a true population estimate. However, recent work at Atka Bay suggests that adult attendance remains stable throughout spring, extending into late November or early December46, so the estimate of population in spring is plausibly based upon a constant demographic component. Further, the image acquisition date is used as a parameter in the model to reflect this.
Previous work24,30 has converted the index of area representing penguins into a number of penguin pairs, using aerial survey observation to ground truth satellite imagery; effectively equating one square metre to a single penguin based on a linear regression of the relationship between satellite derived area and actual counts from aerial survey24, or via a parameter in the Bayesian model30. These population estimates then reflect spring adult attendance at the colony, rather than a full breeding population estimate. In our analysis we are concerned with relative change, rather than population size per sea. Thus, we remove this potential source of error by reporting only the area of penguins as an index of abundance.
Interpretation of results
Our results build on those of LaRue et al.30 and show an evident decline in population numbers in our sector of interest; this study suggests a decline in emperor penguin populations with a best fit of −22% over the 15 year period, equivalent to an absolute decline of 1.6 % per year (N15 = (100 − 22) × N0 = λ15 × N0, which gives λ = 0.984, meaning an annual decline of ca. 1.6%) or a log-linear annual rate of change of −2.25% per year. We report this decline with a very high probability (0.91 for a 30% decline over three generations (a total of 48 years as each generation is 16 years on average; cf 16.4 years reported by Bird et al.47). Our results indicate a steep decline in population until 2016, followed by an undulating plateau in population size after that point. Although the 15-period of our study is relatively short for a long-lived species, it does approximately represent one breeding lifetime of the emperor penguin, which on average lives 20 years and first breeds at around 5 years of age. As indicated above, successful annual breeding for this species is more critical than most other birds and skipping breeding in poorer years is not a long-term option. Therefore, the relatively short time series shown here is important and highly relevant to our study that it might be for many other avian species.
Our analyses reflect a quarter of the Antarctic coastline and, as such, may not be representative of the whole continent. Environmental conditions and patterns of sea-ice extent are regional (Fig. 4). Our sector of interest has been subject to a greater decline in sea-ice extent, especially in the Bellingshausen Sea region.
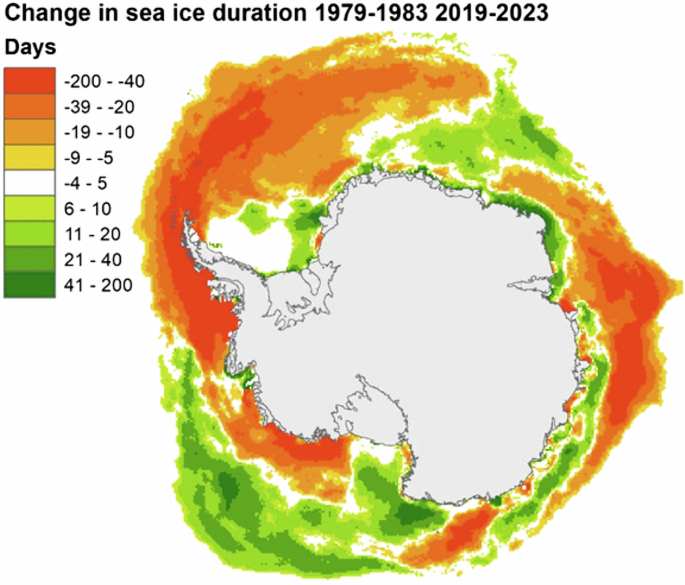
Change in sea ice duration (in days per year) around Antarctica (Sea Ice Index dataset: https://nsidc.org/data/g02135/versions/3 accessed 01/08/2024.
Interestingly, although the sea-ice duration in the Bellingshausen Sea sector has reduced significantly (Fig. 5A), the decrease in our emperor penguin index for this region has not tracked sea-ice decline. This is in marked contrast to the Weddell Sea (Fig. 5B), where sea-ice duration has been variable31, but the decline in our emperor penguin index has been significant. In the Bellingshausen Sea region, sea-ice loss in 2022 was extensive and three of the four colonies analysed in the region in this study had no adults present33. It is plausible that breeding was initiated at these colonies, but breeding failure subsequently occurred. When the sea ice returned in 2023, numbers were lower than average.
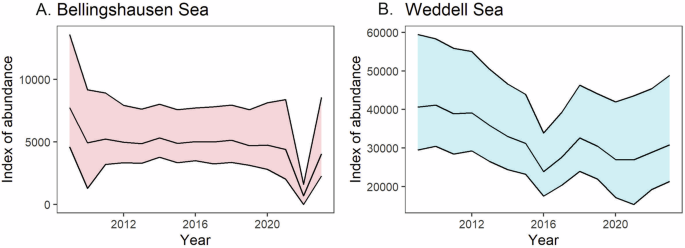
A Bellingshausen Sea regional index of abundance (based on x colonies) showing the extreme event in 2022. B Weddell Sea regional index of abundance (based on y colonies). The loss of the Halley Bay colony in 2016 is evident, followed by a slight upturn as adults relocated to the Dawson-Lambton colony.
In the Weddell Sea, the collapse of the Halley Bay colony in 201634 is a major factor. This colony was originally one of the largest in Antarctica but suffered breeding failures in each year subsequently. After the sea ice break-up in 2016, most of the adult population emigrated to the Dawson Lambton colony, 85 km to the south, where sea ice conditions remained stable. Taken together, the Halley Bay and Dawson Lambton index of abundance showed that the adult population had recovered by 2018 (see Supplementary Fig. 2). The loss of production between 2016 and 2018 is likely to have been significant and may account for a degree of inter-annual variation in the Weddell Sea combined index. Importantly, however, the combined index for the two colonies shows a decline prior to 2016, with an abundance index of ~20,000 in 2010 and ~12,000 in 2015 (Supplementary Figs. 1 and 2). The underlying cause of this decline before 2016 remains unclear.
Comparison with the published results from LaRue et al.30 (see Supplementary Fig. 3), on a regional and overall level, shows good correspondence between 2009 and 2015, with each year’s combined overall value within 7% of each other. However, in the years 2016–2018 our estimates are lower than the LaRue estimates of the same colonies (by 18.6%, 20.4% and 17.8% respectively, see Supplementary Data 6). In 2016 and 2017, this can be partially explained by a slight difference in methodology, specifically of treatment of the zero count at Halley Bay due to early sea ice loss in these years. In our analysis, colonies that have no penguins in the imagery are counted as zero, reflecting the abundance at the time, but in the LaRue et al.30 analysis they are classed as null on the assumption that a breeding population was at the site in the winter before the image was taken in the spring. This can be seen as a difference between our aim to estimate relative springtime abundance, rather than provide an absolute population estimate at each site in each year. However, this reasoning does not fully explain all of the discrepancy in 2016, 2017 and 2018. In these years, lower estimates at three sites: Atka Bay, Gould Bay and Smyley Island are responsible for 88% difference, with the largest difference at Smyley Island (44% of the total difference). Most colonies, however, still show similar estimates in these years. Why these three colonies exhibited very different results is difficult to assess, but it does highlight the high potential variance of estimating emperor population at individual colonies using this methodology.
Why have emperor penguin populations declined?
Attributing a reason why emperor penguin numbers have declined in this sector is complex and, as yet, poorly understood. Previous work has shown that population numbers are affected by fast ice extent, with low fast ice leading to early ice break-out and poor breeding success and excessive fast ice extent leading to higher rates of adult mortality5,48,49. Reduced sea ice extent and early ice-breakout since 2016 have been noted in several areas, but the majority of the population decline shown in this paper has been in the period between 2009 and 2015, before the well documented years of extreme low sea ice that have been observed recently7,8. It must follow, therefore that other environmental factors are the main drivers of population decrease in this sector.
One colony, located within this sector but not included in this study, is the site on the Dion Islands that is likely to be functionally extinct. This colony decreased in population almost continually from the late 1970s until 2009, when no breeding birds were recorded37. Since its discovery in the 1940’s the small colony was located on an island, so breeding success due to lack of sea ice as a breeding platform could not have been a major factor, and other drivers must have reduced the population. The location, in Marguerite Bay on the West Antarctic Peninsula has witnessed rapid warming over the last 50 years50,51 and the Trathan et al. 37 study lists some of the implications of this warming as potential drivers of change. They include reduced prey availability as pack-ice conditions change and oceans warm; increased competition from other predators as they move into the area to exploit the marine resources; lack of fresh snow at the site that emperors use to re-hydrate while breeding; and increased disturbance and predation as other predators move into the area with the warmer conditions and more open ocean environment. It is likely that these factors may now be affecting several of the other emperor colonies in this sector.
The other ice-obligate penguin species, the Adélie (Pygoscelis adeliae), has been shown to have adapted its foraging strategy when sea ice cover is low. In times of no or little sea ice, adults must dive deeper to catch prey, which takes more effort and lowers breeding success52,53. Conversely, and much like earlier studies on emperor penguins, excessive sea ice cover can also be detrimental as adults need to walk further across the ice to access foraging grounds. Recent work has also shown that juvenile emperor penguins exhibit very high interannual variability in body mass, which has important impacts on juvenile survival, and which is tightly associated with sea ice extent during the chick-rearing period12.
There are also other potential drivers linked to rainfall and storminess. Studies on Adélie penguins have shown that increased storms and extreme rainfall can seriously impact breeding success54,55. Extreme storms and precipitation events result in flooding of nests, which drenches chick,s necessitating the use of more energy to regulate temperature. Although nest flooding is not a problem for emperors, personal observations (PTF) support this hypothesis that chick drenching can contribute to mortality as an unusually early extreme rainfall event in mid-October was observed causing the death of several hundred otherwise healthy chicks at the Snow Hill Island colony in 2010. The Antarctic Peninsula has already seen increased storminess, snowfall and rainfall56 and it is likely that, for more northerly or warmer colonies, increased and earlier rainfall events around the coast could be a driver of population change. Other potential causes of chick wetting linked to warming temperatures, such extensive melt pools on the surface of the sea ice in warm years, could also have detrimental effects on breeding success. Analogous to the situation for Adélie chicks, these pools can lead to chick wetting and increased energy expenditure, but like all of these environmental factors, these potential drivers have not yet been quantified for emperor penguins.
A final consideration is huddle size57. As colony size decreases, the huddling ability of emperors in winter or during spring storms becomes less effective. As extreme events are predicted to increase and colonies decrease in size, a smaller huddle size may lead to failed incubation and poor body condition in over-wintering adults.
If we are to understand the drivers of emperor penguin populations and more effectively project their future status it is essential that studies on various potential drivers linked to warming temperatures are implemented. However, what is clear is that emperor penguin populations are already in decline.
Conclusion
The decline of the emperor penguin population in our sector of interest demonstrates a clear negative trend. Although there is significant variance in the model, the best fit results suggest a 22% decline over the 15-year period, something that is much greater than the rate of decline predicted by the process-oriented metapopulation demographic models based on high emission IPCC climate change scenarios (e.g.1) (Fig. 6).
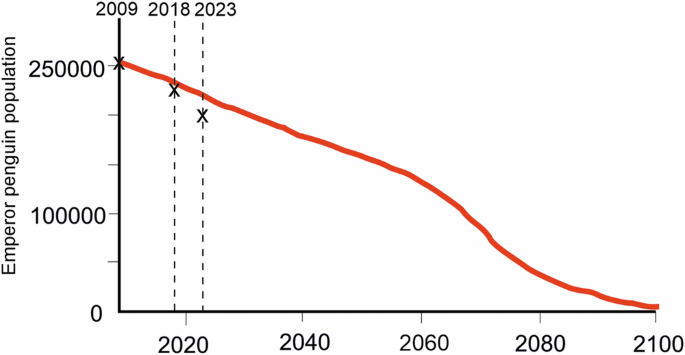
The first X on the left marks the initial global population estimate (Trathan et al.2) of 256,000 breeding pairs, the second denotes the estimated global decline of 9.5% reported by LaRue et al.30 and the third is (best fit) decline of 22% reported in this paper, albeit expanded from a sectorial scale analysis; comparing the results from this regional assessment to the modelled global projection.
Our results support the findings of LaRue et al.30; however, we also show that populations in our sector of study continued to decline after 2018. Our overall results indicate a 91% probability of a 30% decline in the adult population in the study area over three generations (~48 years). Should this trend be reflected at a circumpolar scale, it suggests that emperor penguins should be classified on the IUCN Red List at “Vulnerable�. Such a change would facilitate designation by the ATCM as an Antarctic Specially Protected Species. We recommend that comparable analyses are now undertaken for other parts of the Antarctic continent.
Data availability
Supplementary Files include three figures and six tables cited as Supplementary Data in the text. The first four tables include results and statistics from Bayesian model. The fifth table displays underlying data including Supplementary Information on satellite imagery and image quality and results of multivariate classification that populate the model. The sixth table gives the full results from the Bayesian model for each colony for each year. These data are also available from UK Polar Data Centre: https://doi.org/10.5285/c8d8ffe6-0aff-493c-b40f-e63f0c35f081. The satellite imagery used in these analyses has only been licensed for single use. Copyright for this underlying imagery is held by MAXAR technologies and is commercially available from https://discover.maxar.com/. Details of the images used are contained in the Supplementary Material.
References
-
Jenouvrier, S. et al. The call of the emperor penguin: legal responses to species threatened by climate change. Glob. Change Biol. 27, 5008–5029 (2021).
-
Trathan, P. N. et al. The emperor penguin—vulnerable to projected rates of warming and sea ice loss. Biol. Conserv. 241, 1–30 (2020).
-
Parkinson, C. L. A 40-y record reveals gradual Antarctic sea ice increases followed by decreases at rates far exceeding the rates seen in the Arctic. Proc. Natl. Acad. Sci. USA 116, 14414–14423 (2019).
-
Fraser et al. Eighteen-year record of circum-Antarctic landfast-sea-ice distribution allows detailed baseline characterisation and reveals trends and variability. The Cryosphere 15, 5061–5077 (2021).
-
Barbraud, C. & Weimerskirch, H. Emperor penguins and climate change. Nature. 411, 183–186 (2001).
-
Meredith, M. et al. Polar regions. in IPCC Special Report on the Ocean and Cryosphere in a Changing Climate 203–320 (Cambridge University Press, 2019).
-
Diamond, R., Sime, L. C., Holmes, C. R. & Schroeder, D. CMIP6 models rarely simulate Antarctic winter sea-ice anomalies as large as observed in 2023. Geophys. Res. Lett. 51, e2024GL109265 (2024).
-
Ionita, M. Large-scale drivers of the exceptionally low winter Antarctic sea ice extent in 2023. Front. Earth Sci. 12, 1333706 (2024).
-
Gilbert, E. & Holmes, C. 2023’s Antarctic sea ice extent is the lowest on record. Weather 79, 46–51 (2024).
-
Labrousse, S. et al. Where to live? Landfast sea ice shapes emperor penguin habitat around Antarctica. Sci. Adv. 9, eadg8340 (2023).
-
Jenouvrier, S., Barbraud, C. & Weimerskirch, H. Long term contrasted responses to climate of two Antarctic Seabird species. Ecology 86, 2889–2903 (2005).
-
Le Scornec, E. et al. Intrinsic and extrinsic drivers of juvenile survival in emperor penguins. Polar Biol. 48, 41 (2025).
-
Antarctic Treaty Consultative Meeting: United Kingdom 2022. Report of the CEP Intersessional Contact Group established to develop a Specially Protected Species Action Plan for the emperor penguin. Working Paper 34. Antarctic Treaty Consultative Meeting XLIV, Berlin, Germany (2022). https://documents.ats.aq/ATCM44/wp/ATCM44_wp034_e.docx; https://documents.ats.aq/ATCM44/att/ATCM44_att067_e.docx (2022).
-
Williams, T. D. The Penguins (Oxford University Press, 1995).
-
Stonehouse, B. Breeding behaviour of the emperor penguin. Nature 169, 760 (1952).
-
Prevost, J. Formation des couples, ponte et incubation chez le manchot empereur. Alauda 21, 141–156 (1953).
-
Prevost, J. ðcologie du Manchot Empereur Aptenodytes forsteri Gray. Herman, Paris (Expeditions polaires francaises, 1961).
-
Gauthier-Clerc, M. et al. Penguin fathers preserve food for their chicks. Nature 408, 928–929 (2000).
-
Jackson, S. & Wilson, R. P. The potential costs of flipper-bands to penguins. Funct. Ecol. 16, 141–148 (2002).
-
Cressey, D. Band of brothers. Nature https://doi.org/10.1038/news.2011.15 (2011).
-
Ancel, et al. Emperors in hiding: when ice-breakers and satellites complement each other in Antarctic exploration. PLoS ONE 9, e100404 (2014).
-
Barber-Meyer, S. M., Kooyman, G. L., Ponganis, P. J. Trends in western Ross Sea emperor penguin chick abundances and their relationship with climate. Antarct. Sci. 20, 3–11 (2008).
-
Fretwell, P. T. & Trathan, P. N. Penguins from space: faecal stains reveal the location of emperor penguin colonies. Glob. Ecol. Biogeogr. 18, 543–552 (2009).
-
Fretwell, P. T. et al. An emperor penguin population estimate: The first global, synoptic survey of a species from space. PLOS ONE 7, e33751 (2012).
-
Frederiksen, M. et al. Spatial variation in vital rates and population growth of thick-billed murres in the Atlantic Arctic. Mar. Ecol. Prog. Ser. 672, 1–13 (2021).
-
Descamps, S. & RamÃrez, F. Species and spatial variation in the effects of sea ice on Arctic seabird populations. Divers. Distrib. 27, 2204–2217 (2021).
-
LaRue, M., Kooyman, G., Lynch, H. & Fretwell, P. Emigration in emperor penguins: implications for interpretation of long-term studies. Ecography 38, 114–120 (2015).
-
Fretwell, P. T. & Trathan, P. N. Discovery of new colonies by Sentinel2 reveals good and bad news for emperor penguins. Remote Sens. Ecol. Conserv. 7, 139–153 (2021).
-
US Fish and Wildlife Service report SPECIES STATUS ASSESSMENT EMPEROR PENGUIN (APTENODYTES FORSTERI). Version1. Falls Church, Virginia https://downloads.regulations.gov/FWS-HQ-ES-2021-0043-0003/content.pdf&ved=2ahUKEwjHu8jmh8-IAxX8WEEAHRruAtkQFnoECAkQAQ&usg=AOvVaw20YWb5HLeqea0ndWWhGIFb (2021).
-
LaRue, M. et al. Advances in remote sensing of emperor penguins: first multi-year time series documenting trends in the global population. Proc. Biol. Sci. 291, 20232067 (2018).
-
Turner, J. et al. Absence of 21st century warming on Antarctic Peninsula consistent with natural variability. Nature 535, 7612 (2016).
-
Fretwell, P. T. A 6 year assessment of low sea-ice impacts on emperor penguins. Antarct. Sci. 36, 3–5 (2024).
-
Fretwell, P. T., Boutet, A. & Ratcliffe, N. Record low 2022 Antarctic sea ice led to catastrophic breeding failure of emperor penguins. Commun. Earth Environ. 4, 273 (2023).
-
Fretwell, P. T. & Trathan, P. N. Emperors on thin ice: three years of breeding failure at Halley Bay. Antarct. Sci. 31, 133–138 (2019).
-
Turner, J., Overland, J. E. & Walsh, J. E. An Arctic and Antarctic perspective on recent climate change. Int. J. Climatol. 27, 277–293 (2007).
-
Turner, J. et al. Record low Antarctic sea ice cover in February 2022. Geophys. Res. Lett. 49, e2022GL098904 (2022).
-
Trathan, P. N., Fretwell, P. T. & Stonehouse, B. First recorded loss of an emperor penguin colony in the recent period of Antarctic regional warming: implications for other colonies. PLoS ONE 6, e14738 (2011).
-
Auger-Methe, M. et al. A guide to state-space modeling of ecological time series. Ecol. Monogr. 91, e01470(2021).
-
Jombart, T. & Ahmed, I. adegenet 1.3-1: new tools for the analysis of genome-wide SNP data. Bioinformatics 27, 3070–3071 (2011).
-
Jombart, T., Devillard, S. & Balloux, F. Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genet.11, 94 (2010).
-
Plummer, M. JAGS: A Program for Analysis of Bayesian Graphical Models Using Gibbs Sampling. In Proc. International Workshop on Distributed Statistical Computing (DSC 2003) (Scientific Research, 2003).
-
R. Core Team R: A language and environment for statistical computing. In R Foundation for Statistical Computing, Vienna, Austria, URL https://www.R-project.org/. https://www.R-project.org/ (2024).
-
Kellner, K. jagsUI: A Wrapper Around ‘rjags’ to Streamline ‘JAGS’ Analyses. R package version 1.5.2, https://CRAN.R-project.org/package=jagsUI (2021).
-
Gelman, A. & Shirley, K. Inference from simulations and monitoring convergence. In Handbook of Markov Chain Monte Carlo (eds Brooks, S., Gelman, A., Jones, G. & Meng, X.-L.) 1st edn, pp. 163–174 (Chapman and Hall/CRC, 2011).
-
Labrousse, S. et al. Quantifying the causes and consequences of variation in satellite-derived population indices: a case study of emperor penguins. Remote Sens. Ecol. Conserv. 8, 151–165 (2022).
-
Winterl, A. et al. Remote sensing of emperor penguin abundance and breeding success. Nat. Commun. 15, 4419 (2024).
-
Bird, J. P. et al. Generation lengths of the world’s birds and their implications for extinction risk. Conserv. Biol. 40, 1252–1261 (2020).
-
Jenouvrier, S. et al. Living with uncertainty: using multi-model large ensembles to assess emperor penguin extinction risk for the IUCN red list. Biol. Conserv. 305, 11037(2025).
-
Jenouvrier, S., Barbraud, C., Weimerskirch, H. & Caswell, H. Limitation of population recovery: a stochastic approach to the case of the emperor penguin. Oikos 118, 1292–1298 (2009).
-
Vaughan, D. et al. Recent rapid climate warming on the Antarctic Peninsula. Clim. Change 60, 243–274 (2003).
-
Meredith, M. & King, J. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Lett. 32, L19604 (2005).
-
Le Guen, C. et al. Reproductive performance and diving behaviour share a common sea-ice concentration optimum in Adélie penguins (Pygoscelis adeliae). Glob. Change Biol. 11, 5304–5317 (2018).
-
Kokubun, N. et al. Abundance and estimated food consumption of seabirds in the pelagic ecosystem in the eastern Indian sector of the Southern Ocean. Prog. Oceanogr. 231, 103385 (2024).
-
Fraser, W. A nonmarine source of variability in Adélie penguin demography. Oceanography 26, 207–209 (2013).
-
Youngflesh, C. et al. Large-scale assessment of intra- and inter-annual breeding success using a remote camera network. Remote Sens. Ecol. Conserv. 7, 97–108 (2021).
-
Vignon, É. et al. Present and future of rainfall in Antarctica. Geophys. Res. Lett. 48, e2020GL092281 (2021).
-
Zitterbart, D. P., Wienecke, B., Butler, J. P. & Fabry, B. Coordinated movements prevent jamming in an emperor penguin huddle. PLoS ONE 6, e20260 (2011).
Acknowledgements
We would like to acknowledge the funding for satellite imagery and analysis time from WWF UK (GB095701), project NEB2181 ‘Understanding emperor penguin populations in the Weddell Sea and Antarctic Peninsula’ and previous WWF funding over the 15-year period. We would also like to thank Jennifer Brown for her help with the early analysis and Norman Ratcliffe for his comments on the manuscript. This paper is linked to the Southern Ocean Observing System’s Censusing Animal Populations from Space (CAPS) Special Action Group.
Author information
Authors and Affiliations
Contributions
Peter Fretwell instigated and designed the project, acquired data and led the image analysis, led the writing of the paper and constructed figures. Jaume Forcada did the statistical analysis, constructed figures and helped write the paper. Connor Bamford did image analysis and helped write the paper. Aliaksandra Skachkova did image analysis, acquired imagery and helped write the paper. All authors helped with edits and comments. Phil Trathan was instrumental in the initial years of data acquisition and led the previous study, which acquired much of the data. He provided input on initial project design and helped write the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Sébastien Descamps and Malgorzata Korczak-Abshire, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Heike Langenberg and Aliénor Lavergne. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article
Cite this article
Fretwell, P.T., Bamford, C., Skachkova, A. et al. Regional emperor penguin population declines exceed modelled projections.
Commun Earth Environ 6, 436 (2025). https://doi.org/10.1038/s43247-025-02345-7
-
Received: 07 November 2024
-
Accepted: 01 May 2025
-
Published: 10 June 2025
-
DOI: https://doi.org/10.1038/s43247-025-02345-7
Search
RECENT PRESS RELEASES
Related Post




{kind=link}
{kind=link}
{kind=link}
{kind=link}